

A polipose nasal é uma doença inflamatória progressiva que reduz a qualidade de vida dos pacientes. Os efeitos das vias apoptóticas e autofágicas na patogênese da polipose nasal ainda não são claramente conhecidos.
ObjetivoInvestigar as vias apoptóticas MAPK/JNK, antiapoptóticas PI3K/mTOR e autofágicas LC3 as quais se encontram relacionadas entre si, nos tecidos da polipose nasal.
MétodoVinte pacientes com pólipos nasais e 15 pacientes submetidos a redução de conchas nasais inferiores foram incluídos. Pacientes com asma, tríade de Samter e sinusite fúngica alérgica foram excluídos. As vias apoptóticas e autofágicas foram investigadas em cortes de tecido nasal embebidos em parafina de 20 amostras de polipose nasal e 15 de amostras provenientes de redução de conchas nasais inferiores com H&E e imuno‐histoquímica com h‐escore. O método TUNEL com índice apoptótico foi usado para demonstrar células apoptóticas.
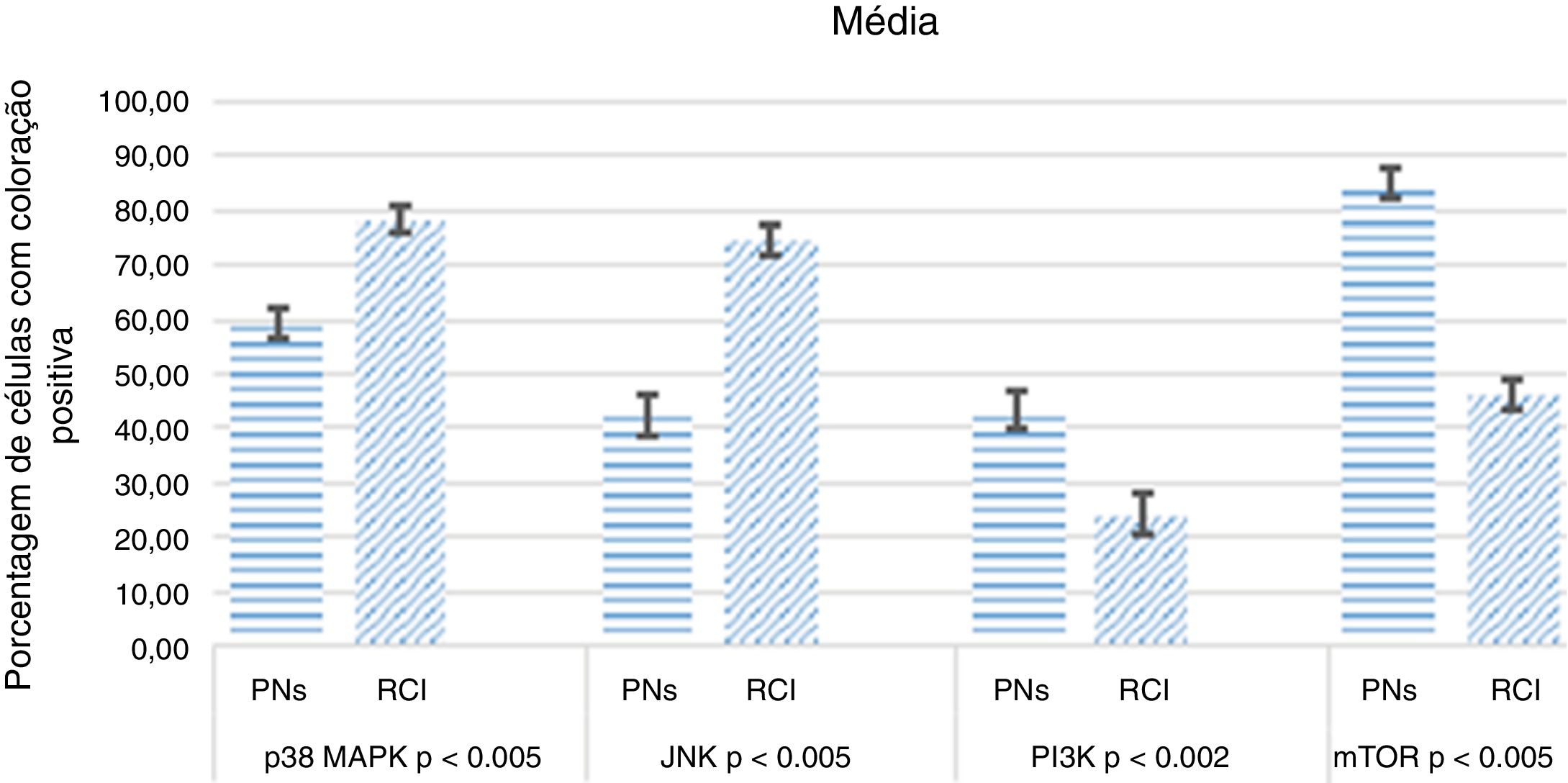
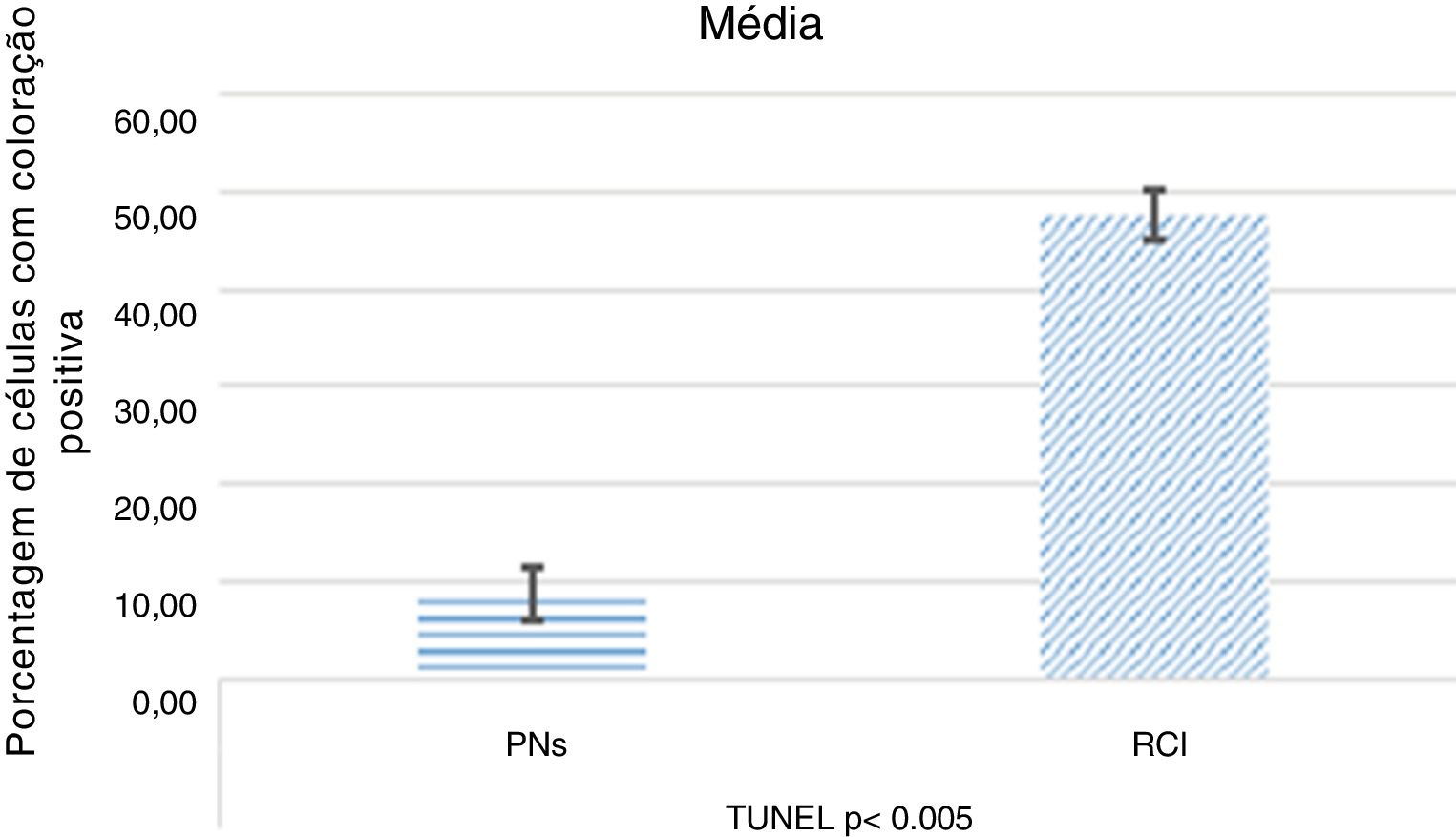
ResultadosImunorreatividade diminuída de P38 MAPK (p <0,005) e JNK (p <0,005) foi observada na polipose nasal comparada à de tecidos oriundos de redução de conchas nasais inferiores. Essa diminuição pode indicar uma down‐regulação da apoptose, como demonstrado pela diminuição da coloração de TUNEL na polipose nasal (p <0,005). A imunorreatividade de PI3K (p <0,002) e mTOR (p <0,005) estava aumentada na polipose nasal. Esse aumento indica uma down‐regulação da autofagia, como demonstrado pela diminuição da coloração de LC3 na polipose nasal (p <0,001).
ConclusãoApoptose e autofagia deficientes através das vias MAPK/JNK e PI3K/mTOR podem ter um papel na patogênese da polipose nasal.
Os pólipos nasais (PNs), lesões benignas que surgem principalmente nas mucosas do nariz e dos seios paranasais, são uma doença comum, com alta taxa de recorrência.1 Apesar da morbidade significativa dessa doença recorrente, os mecanismos centrais da patogênese da polipose nasossinusal são complexos e mal compreendidos.
Supõe‐se que os PNs sejam caracterizados por alterações morfológicas, como hiperplasia e metaplasia do revestimento epitelial, infiltração de células inflamatórias e fibrose estromal e edema.2 Acredita‐se que muitas citocinas e quimiocinas tenham uma grande contribuição na patogênese dos PNs.3 Por exemplo, os eosinófilos são as células inflamatórias mais comuns.4 Pesquisas anteriores demonstraram que o aumento da síntese de colágeno devido à inibição da apoptose dos eosinófilos é um fator que contribui para o desenvolvimento de pólipos.5 Além disso, alguns pesquisadores relataram que a apoptose nas células inflamatórias é um fator importante na resolução dos PNs.6 Portanto, a apoptose tem um papel na remoção de células indesejadas associadas aos PNs. Além disso, estudos recentes sugeriram que a apoptose celular tardia tem um papel importante na patogênese dos PNs.7 Nesse sentido, mostrar a via apoptótica verdadeira pode ajudar a impedir a formação de PNs.
Por outro lado, a autofagia é um processo altamente específico para mitigar vários tipos de estresse celular. Nesse processo, o conteúdo citoplasmático é sequestrado, transportado por autofagossomo de membrana dupla para os lisossomos e degradado. Várias vias são responsáveis por regular a apoptose e a autofagia. As proteínas quinases ativadas por mitogênio (MAPKs, do inglês Mitogen‐Activated Protein Kinases) desempenham um papel importante na regulação de muitos processos celulares, inclusive proliferação, diferenciação e apoptose celular. As MAPKs consistem em MAPKs ativadas por estresse, quinases de c‐Jun NH2‐terminal (JNKs) e MAP38 p38,8 o que significa que uma baixa atividade de MAPK/JNK suprime a apoptose. Por outro lado, as vias MAPK, a via de transdução de sinal fosfatidilinositol 3‐quinase/alvo da rapamicina,P13K/mT, em mamíferos, uma molécula sinalizadora para síntese de proteínas e sobrevivência celular são ativadas e inibem a autofagia,9 o que significa que uma alta atividade de PI3K/mTOR suprime a autofagia.
Também foi relatado que a molécula LC3 interage com a via mTOR na formação da membrana autofagossômica.10 A LC3‐II está localizado na membrana dos autofagossomos quando a autofagia é ativada e a LC3‐II aumenta diretamente proporcional ao número de autofagossomos.11
A autofagia e a apoptose são os importantes reguladores do destino celular. Essa regulação as torna importantes na doença inflamatória. Não há consenso sobre o papel desses dois mecanismos na fisiopatologia dos PNs. O objetivo deste artigo é investigar o papel das vias apoptótica e relacionada à autofagia, como MAPK/JNK, PI3K/mTOR e LC3 na fisiopatologia dos PNs.
MétodoPacientes e tecidosO estudo foi aprovado pelo Izmir Katip Celebi University Atatürk Education and Research Hospital. O consentimento informado foi obtido de cada paciente e o estudo foi aprovado pelo comitê de ética da Izmir Katip Celebi University sob o número 192/13. O estudo incluiu 20 pacientes com PN e 15 tecidos de concha nasal (TCN) e a faixa etária foi de 18 a 55 anos. Além disso, pacientes com sinusite fúngica alérgica, asma, intolerância ao AAS e tríade de Samter não foram incluídos no estudo. Os pacientes recrutados não usaram esteroides sistêmicos ou tópicos, nem tomaram outro medicamento sistêmico por pelo menos quatro semanas antes da cirurgia. Amostras de PNs foram coletadas de pacientes submetidos a biópsia de pólipo por via endoscópica para fins de diagnóstico e amostras de tecido de conchas nasais (TCN) 1,3,12,13 foram coletadas de pacientes submetidos a ressecção das conchas nasais devido à a hipertrofia compensatória de conchas secundária a desvio do septo nasal.
H&E e imuno‐histoquímicaTodas as amostras foram fixadas em formalina a 10% por 24 horas e processadas para ser embebidas em parafina, com o protocolo de rotina. Cortes de 5μm de espessura foram obtidos e corados com hematoxilina e eosina (H&E). Também foram usados cortes adicionais para coloração imuno‐histoquímica. As amostras de tecido foram armazenadas a 60°C durante a noite e depois foram desparafinadas com xileno por 30 minutos. Os tecidos foram então tratados com 2% de tripsina e incubados em solução de H2O2 a 3%. Em seguida, os cortes foram incubados com anticorpos primer anti‐p38MAPK (sc‐7973, Santa Cruz Biotechnology, Inc.), anti‐JNK (sc‐7345, Santa Cruz Biotechnology, Inc.), anti‐PI3K (sc‐1637, Santa Cruz Biotechnology, Inc.), anti‐mTOR (sc‐8319, Santa Cruz Biotechnology, Inc.) e LC3 (LC3B, NB100‐2220, Novus Biologicals, Littleton, CO, EUA) em uma diluição de 1/100 por 18 horas a +4°C. Após a administração do anticorpo secundário, os cortes foram corados com o sistema de substrato DAB que continha diaminobenzidina (DAB‐plus substrate kit, Invitrogen) para detectar a imunorreatividade. Eles foram examinados de forma cega por microscopia óptica (Olympus BX‐43, Tóquio, Japão).
Duas áreas selecionadas aleatoriamente foram escolhidas e, nas secções em que toda a coloração parecia intensa, um campo aleatório foi escolhido. Foram marcadas pelo menos 100 células por campo X40 para cada tecido em todos os grupos. Todas as secções foram escolhidas de maneira semiquantitativa, consideraram‐se a intensidade e a porcentagem de coloração celular. As intensidades foram classificadas em leve (1+), moderada (2+), forte (3+) e muito forte (4+). A coloração dos antibióticos primer foi graduada de maneira semiquantitativa e o escore H foi calculado com a seguinte equação: Escore H=ΣPi (i+1), onde i=intensidade da coloração com valores 1, 2, 3 e 4 (leve, moderado ou forte, muito forte) e Pi é a porcentagem de células coradas para cada intensidade, varia de 0 a 100.14,15
Método TUNELPara distribuição das células apoptóticas, foi usada a técnica de Terminal Deoxynucleotidyl Transferase Mediated dUTP Nick End Labeling (TUNEL), de acordo com o procedimento do fabricante do kit (S7101, ApopTag, Millipore). Após todas as etapas, a montagem foi feita com Entellan e elas foram examinadas independentemente por dois histologistas e as células, TUNEL‐positivas ou não, foram contadas em campos escolhidos aleatoriamente para cada caso. Foi determinado o índice apoptótico como a porcentagem de células TUNEL‐positivas com imunocoloração marrom positiva. Dois observadores cegos para a atribuição do tratamento avaliaram as pontuações de coloração de forma independente.
Análise estatísticaA homogeneidade dos dados foi avaliada pelo teste de Shapiro‐Wilk. Para dados normalmente distribuídos, foi usado o teste t de Student; caso contrário, usamos o teste ×2 ou o teste U de Mann‐Whitney. O software SPSS (versão 16) foi usado para todas as análises estatísticas. O nível de significância estatística foi estabelecido em p=0,05 e foram determinados intervalos de confiança de 95%.
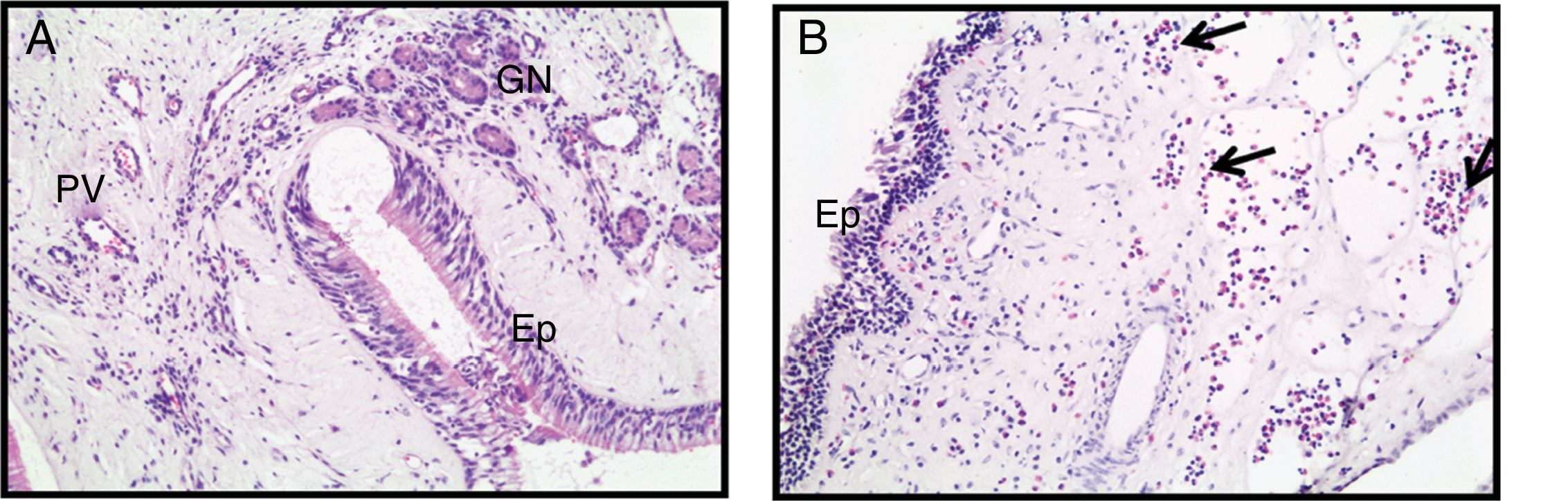
ResultadosHistologia dos tecidos nasaisAs amostras de tecidos nasais coradas com hematoxilina e eosina são apresentadas na figura 1. Nas amostras de tecido de conchas, o plexo venoso submucoso e as glândulas nasais são demonstradas (fig. 1A). A formação de PNs é caracterizada histologicamente por edema submucoso e acúmulo de matriz extracelular (fig. 1B). Além disso, nos PNs, a infiltração celular na submucosa, especialmente as células eosinófilas, foi significativamente maior do que no tecido de concha nasal, onde foram observadas lesões epiteliais (fig. 1B).
Avaliação imuno‐histoquímica – Apoptótica e autofágica e PN (B). (Ep, epitelial; PV, plexo venoso; GN, hlândulas nasais; Seta, eosinófilos) (Ampliação original × 200).")
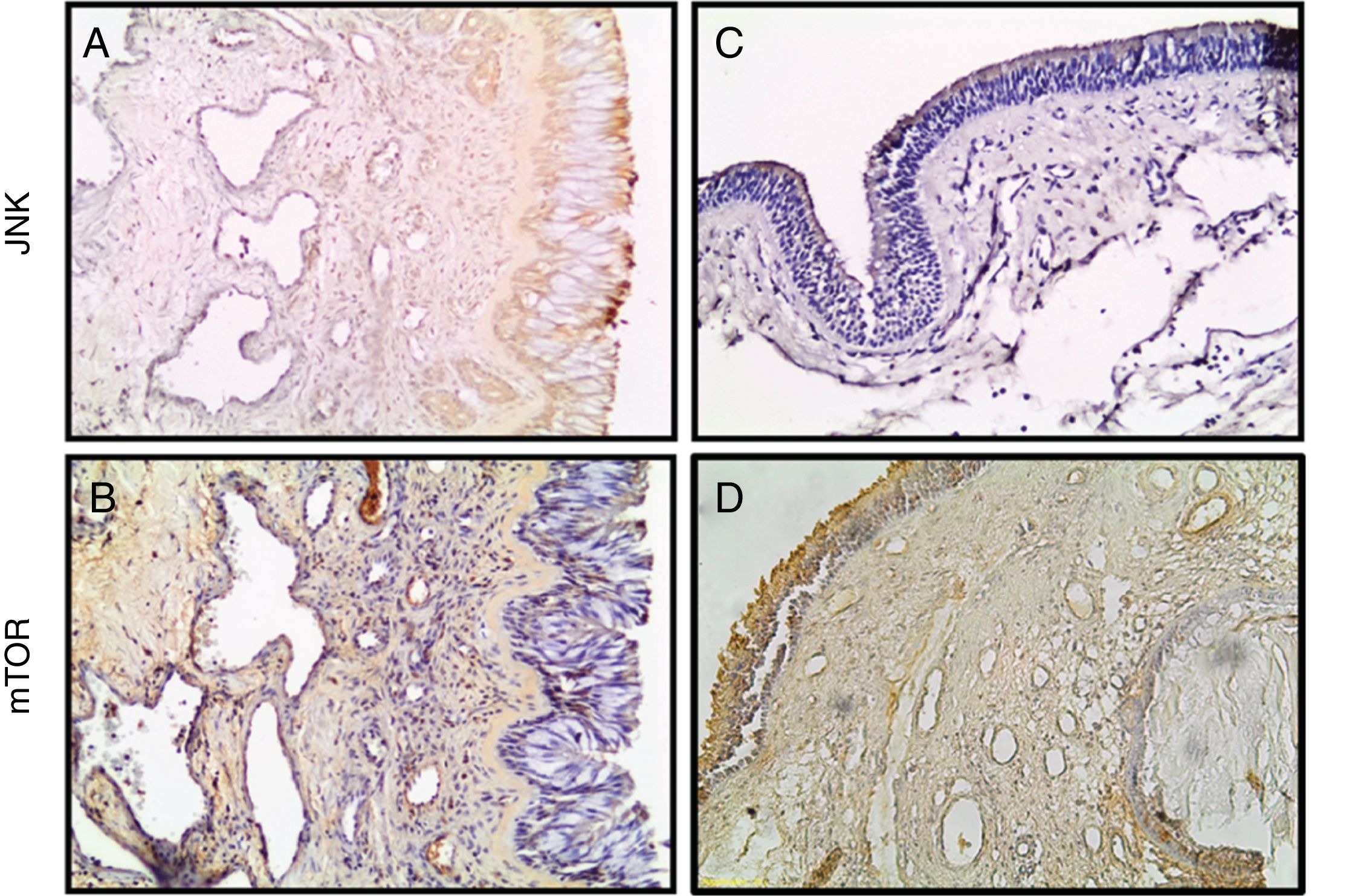
Estudos imuno‐histoquímicos em 35 secções adicionais de tecido nasal embebido em parafina revelaram a localização dos marcadores relacionados à apoptose e a relação entre a expressão proteica e os tecidos de PNs.
As imunorreatividades positivas foram determinadas nas partes epitelial e estromal de todos os tecidos. Exemplos de coloração imuno‐histoquímica são mostrados na figura 2. A imunorreatividade anti‐JNK foi diminuída e a imunorreatividade mTOR estava aumentada em PNs (fig. 2).
 e PNs (C, D). (Ampliação original × 200).")
Para o anticorpo primário P38 MAPK, a coloração imuno‐histoquímica foi moderada e imunorreatividades leves a moderadas de anti‐PI3K foram encontradas nos PNs.
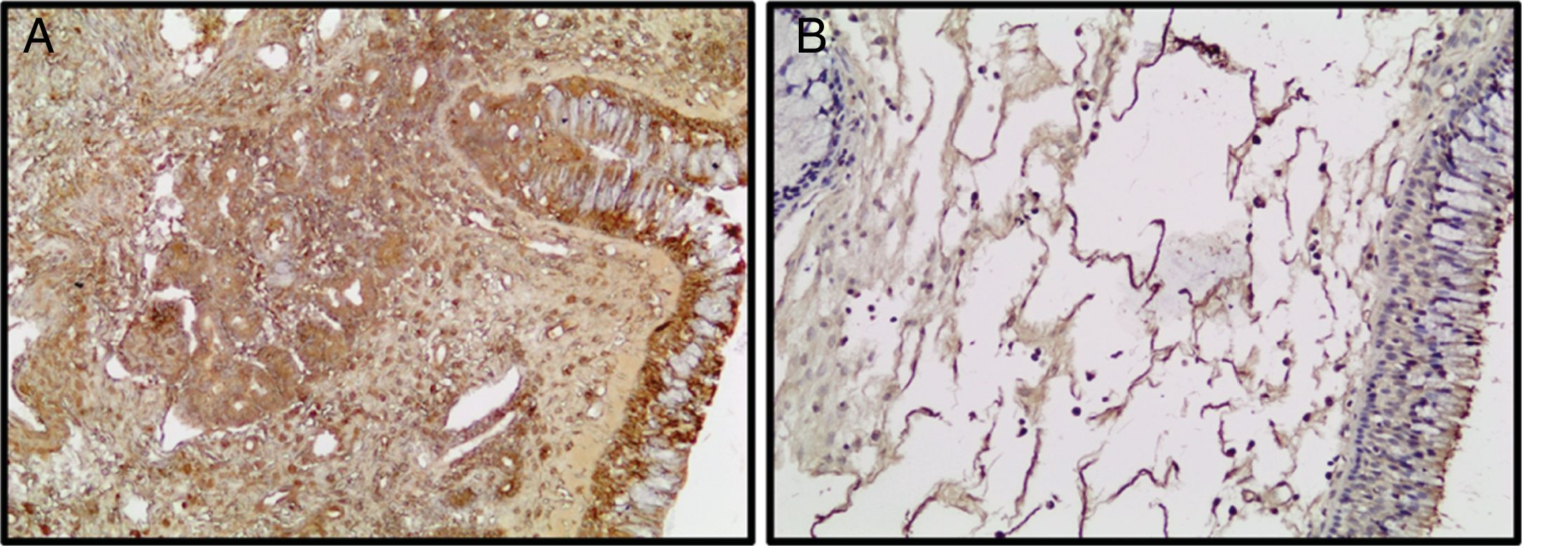
Um marcador autofágico, a imunorreatividade de LC3 estava diminuída nos PNs em relação ao grupo controle (fig. 3).
 e PNs (B). (Ampliação original × 200).")
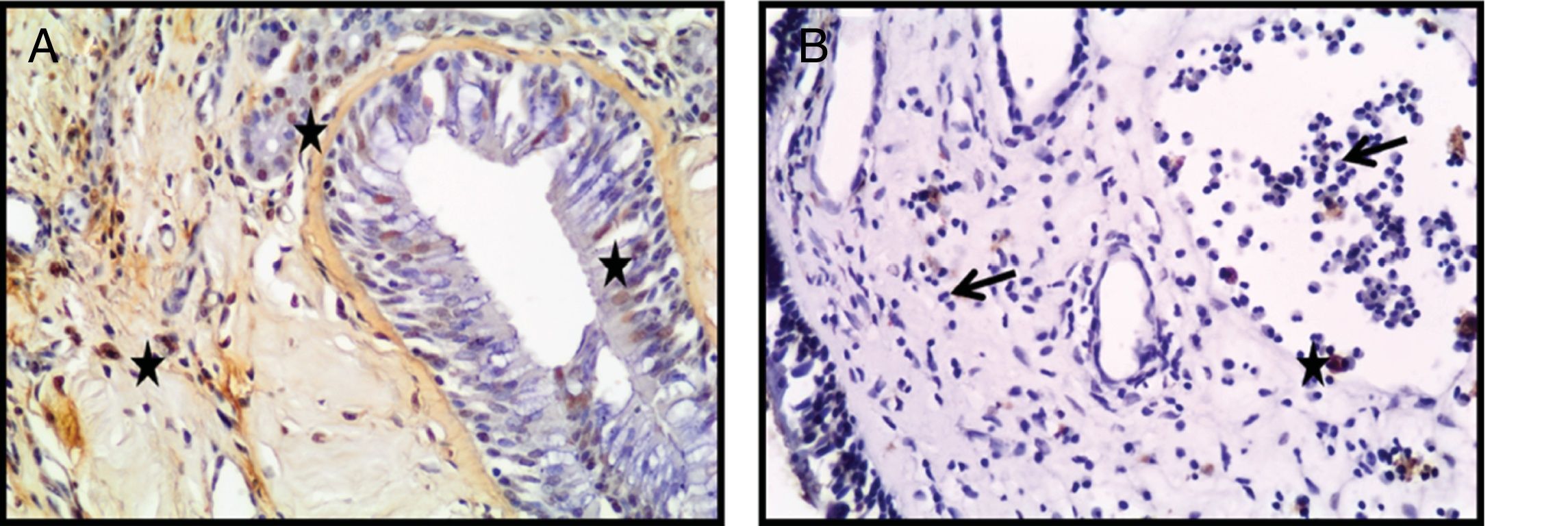
Como resultado da coloração do método Terminal Deoxynucleotidyl Transferase Mediated dUTP Nick End Labeling (TUNEL), para avaliar a presença de células apoptóticas e fragmentações de DNA nuclear nos pacientes, a média de células positivas para TUNEL encontradas foi moderada para tecido de concha nasal (fig. 4A) e negativa a fraca para PNs (fig. 4B). Além disso, detectamos que, além de o número das células apoptóticas ser menor nos tecidos de PNs, a apoptose não foi encontrada nos eosinófilos ou nas outras células inflamatórias (fig. 4B). figura 5, figura 6
 e PNs (B). As células TUNEL‐positivas foram identificadas pela coloração marrom (estrela) (Seta: eosinófilos) (A, B; ampliação original × 400).")
 e tecido de redução de concha inferior (RCI).")
 e amostras de redução de concha inferior (RCI).")
As intensidades de coloração positiva de P38 MAPK, JNK, PI3K, mTOR, LC3 e TUNEL no TCN e nos tecidos nasais estão resumidas na tabela 1.
Porcentagem de células positivas para coloração em pólipos nasais (PNs) e tecido de conchas nasais (TCN)
| P38 Mapk | JNK | PI3K | mTOR | LC3 | Tunel | |
|---|---|---|---|---|---|---|
| PNs | Moderadop ˂ 0,005 | Fraco/Moderadop ˂ 0,005 | Leve/Moderadop ˂ 0,002 | Fortep ˂ 0,005 | Fortep ˂ 0,001 | Negativo/Fracop ˂ 0,005 |
| TCN | Forte | Forte | Negativo | Moderado | Leve/Moderado | Moderado |
Os resultados da análise mediana da coloração estão resumidos nas figuras 5 e 6.
DiscussãoNeste estudo, demonstramos que a diminuição das expressões das vias P38/MAPK, JNK e LC3 e o aumento da expressão das vias PI3K e mTOR podem estar relacionados à down‐regulação da apoptose e autofagia nos PNs.
A apoptose é a morte celular programada, na qual as células inúteis, danificadas irreparavelmente ou indesejadas são eliminadas durante o desenvolvimento.16 A autofagia é um processo fisiológico normal no corpo envolvido na homeostase celular e nos mecanismos de sobrevivência nas células com respiração normal.17 Embora se saiba pouco sobre o papel da autofagia em doenças respiratórias, como rinite alérgica e sinusite,18 não há consenso na literatura sobre o papel da autofagia e apoptose na fisiopatologia dos PNs.
Chen et al. estudaram a expressão da proteína Light Chain 3 (LC3), um indicador comum da autofagia, e a expressão da proteína quinase B – alvo mecanístico da rapamicina (Akt‐mTOR) em amostras de tecido fresco de 5 pólipos nasais e 6 mucosas nasais normais.3 Eles observaram que a expressão de LC3 estava diminuída e a atividade de Akt‐mTOR aumentada em PNs. Wang et al. também relataram diminuição da autofagia.19 Em seu estudo, eles verificaram que a expressão de LC3 estava diminuída, enquanto a expressão de COX‐2 estava significantemente aumentada em tecidos frescos de PNs em comparação com o controle de mucosa normal. A autofagia era deficiente nos tecidos de PNs e a COX‐2 era regulada negativamente pela autofagia nos fibroblastos derivados de PNs.
Em outro estudo, Qi et al. sugeriram que as expressões de Beclin1 e LC3 nos tecidos de pólipo nasal eram menores do que a expressão da mucosa de concha inferior.20
Ao contrário, Shun et al. relataram que a expressão de HIF‐1α, LDH e LC3II estava aumentada em amostras de pólipos nasais. Eles concluíram que a análise de seus dados indicava que a hipóxia pode contribuir para a formação do PNs ao promover a autofagia nos fibroblastos dos PNs.21 Wang et al. relataram que a expressão das proteínas autofágicas LC3 estava aumentada nos tecidos dos pólipos e a via de sinalização da apoptose extrínseca era notavelmente ativada nas células epiteliais de PNs.22
Küpper et al. investigaram a expressão do mRNA dos mediadores da apoptose, como caspase 3, 7 e 9, e da proteína p53 analisada com a reação em cadeia da polimerase via transcriptase reversa quantitativa em 25 PNs e 18 amostras de controle. Eles relataram a expressão reduzida de p53 com caspase 3 e 9 em pacientes com RSCcPNs (rinossinusite crônica com PNs) comparados aos dos controles. Eles concluíram que a expressão reduzida desses fatores de apoptose na RSCcPNs pode estar relacionada à maior proliferação e à perpetuação de células inflamatórias.23 Em nosso estudo, observamos apoptose e autofagia diminuídas em amostras de PNs em comparação com amostras de TCN.
Por outro lado, em anos anteriores, houve aumento do número de estudos que mostraram o papel do estresse oxidativo na patogênese da polipose nasal.24 MAPKs é a principal via de transdução de sinal sensível ao estresse oxidativo e intracelular.25 Portanto, a expressão de MAPK pode ser um dos principais determinantes da apoptose nos PNs.
Além da imuno‐histoquímica, investigamos as células apoptóticas pelo método TUNEL. A porcentagem de células positivas para TUNEL encontradas estava diminuída nos PNs comparadas ao tecido de concha nasal. Particularmente, não houve apoptose nos eosinófilos nos tecidos de PNs. O eosinófilo é a principal célula responsável pela fisiopatologiados PNs.26 Em situações patológicas, a produção de eosinófilos é aumentada e essas células migram para os tecidos inflamados, nos quais se acredita que sua vida útil aumente através de uma combinação de mediadores inflamatórios e alterações no microambiente local.27 A remoção de eosinófilos do tecido polipoide ocorre através da apoptose.28
Os eosinófilos têm funções citotóxicas, como a liberação de proteínas básicas importantes, a peroxidase eosinofílica e a proteína catiônica eosinofílica, que causam danos aos tecidos e a eosinofilia aumenta a probabilidade de doença recorrente.29 Khalmuratova et al. testaram a wogonina (5,7‐di‐hidroxi‐8‐metoxiflavona) em PNs e sugeriram que a wogonina atenua a formação de pólipos nasais ao induzir apoptose por eosinófilos.30 Em outro estudo, a expressão de PARP e caspase 8 estavam aumentadas nos tecidos de PNs tratado com bleomicina e os tecidos diminuíram e depois desapareceram.31
Esses resultados mostram que o aumento da expressão de moléculas antiapoptóticas diminuiu a expressão de moléculas pró‐apoptóticas e autofágicas nos PNs e, principalmente, a ausência da apoptose nas células eosinofílicas nos PNs pode ser fator contribuinte para o desenvolvimento de pólipos.
LimitaçõesUsamos métodos imuno‐histoquímicos em vez de testes de expressão gênica e western blot, que são quantitativos. Para diminuir a subjetividade dos resultados, dois observadores cegos avaliaram os resultados conforme descrito nos métodos.
ConclusãoApoptose, eosinofilia e hiperproliferação são os principais processos celulares na polipose nasal e essas proteínas podem participar e desempenhar alguns papéis importantes para a obtenção de novas informações sobre essa doença e direcionamento de protocolos de tratamento.
FinanciamentoCoordenação de Projetos e Pesquisa Científica da Izmir Katip Çelebi University (n° 2013‐3‐TSBP‐16).
Conflitos de interesseOs autores declaram não haver conflitos de interesse.
Como citar este artigo: Simsek F, Eren E, Bahceci S, Aladag I. High PI3K/mTOR and low MAPK/JNK activity results in decreased apoptosis and autophagy in nasal polyposis. Braz J Otorhinolaryngol. 2021;87:572–7.
A revisão por pares é da responsabilidade da Associação Brasileira de Otorrinolaringologia e Cirurgia Cérvico‐Facial.
gology tem o prazer em homenagear os revisores


