
Salicylate at high doses induces tinnitus in humans and experimental animals. However, the mechanisms and loci of action of salicylate in inducing tinnitus are still not well known. The expression of Immediate Early Genes (IEG) is traditionally associated with long‐term neuronal modifications but it is still not clear how and where IEGs are activated in animal models of tinnitus.
ObjectivesHere we investigated the expression of c‐fos and Egr‐1, two IEGs, in the Dorsal Cochlear Nucleus (DCN), the Inferior Colliculus (IC), and the Posterior Ventral Cochlear Nucleus (pVCN) of rats.
MethodsRats were treated with doses known to induce tinnitus in rats (300mg/kg i.p. daily, for 3 days), and c‐fos and Egr‐1 protein expressions were analyzed using western blot and immunocytochemistry.
ResultsAfter administration of salicylate, c‐fos protein expression increased significantly in the DCN, pVCN and IC when assayed by western blot. Immunohistochemistry staining showed a more intense labeling of c‐fos in the DCN, pVCN and IC and a significant increase in c‐fos positive nuclei in the pVCN and IC. We did not detect increased Egr‐1 expression in any of these areas.
ConclusionOur data show that a high dose of salicylate activates neurons in the DCN, pVCN and IC. The expression of these genes by high doses of salicylate strongly suggests that plastic changes in these areas are involved in the genesis of tinnitus.
Salicilato em doses elevadas induz zumbido nos seres humanos e em animais experimentais. No entanto, os mecanismos e loci de ação do salicilato na indução de zumbido ainda não são bem conhecidos. A expressão dos genes precoces imediatos (GPIs) está tradicionalmente associada a alterações neuronais em longo prazo, mas ainda não está claro como e onde os GPIs são ativados em modelos animais de zumbido.
ObjetivosNo presente estudo investigamos a expressão de c‐fos e Egr‐1, dois GPIs, no núcleo coclear dorsal (NCD), colículo inferior (CI) e núcleo coclear ventral posterior (NCVp) de ratos.
MétodosOs ratos foram tratados com doses que, conhecidamente, induzem zumbido em ratos (300mg/kg IP/dia, por três dias) e as expressões das proteínas c‐fos e Egr‐1 foram analisadas por meio de Western blot e imunoistoquímica.
ResultadosApós a administração de salicilato, a expressão da proteína c‐fos aumentou significativamente no NCD, NCVp e CI, quando analisados por Western blot. A coloração imunoistoquímica mostrou uma marcação mais intensa de c‐fos no NCD, NCVp e CI e um aumento significativo de núcleos positivos de c‐fos no NCVp e CI. Não detectamos aumento da expressão de Egr‐1 em qualquer dessas áreas.
ConclusãoNossos dados mostram que uma dose alta de salicilato ativa neurônios no NCD, NCVp e CI. A expressão desses genes por doses altas de salicilato sugere que as alterações plásticas nessas áreas estão envolvidas na gênese do zumbido.
O zumbido é uma sensação sonora fantasma que pode ser a consequência de vários fatores, inclusive trauma acústico, drogas, distúrbios mandibulares temporais ou surdez.1 Os mecanismos de indução e manutenção do zumbido ainda são uma questão de debate, especialmente porque o zumbido pode resultar de diferentes condições. Doses elevadas de salicilato induzem zumbido em seres humanos e doses de 150‐400mg/kg induzem sintomas comportamentais de zumbido em animais experimentais em menos de 24 horas e esse efeito desaparece em até 72 horas após a interrupção do tratamento.2
A despeito do uso comum de salicilato como agente indutor de zumbido, seus mecanismos e loci de ação ainda são obscuros. Experimentos com imagens in vivo em ratos demonstraram que altas doses de salicilato causam hiperatividade em áreas auditivas específicas, como o colículo inferior (CI), o núcleo coclear dorsal (NCD) e o córtex auditivo (CA), mas não o núcleo coclear ventral (NCV).3,4 Por outro lado, estudos da expressão do gene precoce imediato c‐fos relataram resultados menos consistentes. Uma dose única de salicilato (350mg/kg) aumentou a expressão de c‐fos apenas no CA de gerbilos.5 Um estudo anterior não mostrou expressão de c‐fos no tronco cerebral auditivo após uma única dose de salicilato, mas apenas em áreas não auditivas, como o locus coeruleus e a área cinzenta periaquedutal.6 Outro estudo observou uma diminuição da expressão de c‐fos no CI de gerbilos.7 Um estudo que usou o tratamento crônico com salicilato (250mg/mL) mostrou um aumento da expressão de c‐fos apenas no CI, e não no NCD.8 A maioria desses dados está de acordo com as observações de que o zumbido induzido por salicilato ativa as vias auditivas extralemniscais, especialmente no NCD.2,3
A expressão dos genes precoces imediatos (GPIs) é considerada um marcador do aumento da atividade cerebral em resposta a diversos estímulos. Esses genes são fatores de transcrição que desencadeiam a expressão de outros genes responsáveis por alterações de longo prazo nos neurônios. A expressão do GPI c‐fos é um marcador comumente usado da atividade neuronal e é rapidamente suprarregulado após a estimulação neuronal.9–12 Já o GPi egr‐1 é ativada em resposta ao influxo de cálcio neuronal e promove alterações funcionais e estruturais nos neurônios, inclusive no sistema auditivo.13,14
Neste estudo, nosso objetivo foi investigar a ativação de c‐fos e egr‐1 no NCD e CI em vias auditivas de ratos submetidos a protocolo de administração de salicilato, que é eficaz na indução de zumbido em ratos (três doses diárias de 300mg/kg).3 Devido à sua proximidade com o NCD, avaliamos também a expressão desses genes no núcleo coclear ventral posterior (NCVp).
MétodoAnimais e tratamento medicamentosoTodos os procedimentos experimentais feitos nos animais foram aprovados pelo Comitê de Ética em Pesquisa Animal da instituição (protocolo n° 011/2013) e seguiram as orientações e recomendações dos Institutos Nacionais de Saúde sobre o tratamento dos animais. As experiências foram feitas em ratos Wistar machos entre 60 e 65 gramas. Os ratos foram agrupados e mantidos em número de 4‐5 por gaiola, em ciclo claro/escuro de 12h/12h, com comida e água ad libitum.
Os animais foram injetados por via IP com 300mg/kg de salicilato de sódio (Sigma) (10μL/g) dissolvidos em soro fisiológico, durante três dias consecutivos, com intervalo de 24 horas para cada dose. Os animais foram, então, anestesiados com isoflurano e sacrificados por decapitação no terceiro dia, três horas após a administração do fármaco. Os grupos controle receberam administração de soro fisiológico de acordo com o mesmo protocolo. Esse protocolo de administração de salicilato demonstrou ser eficaz na indução de zumbido em ratos.3
Detecção de egr‐1 e c‐fos com o uso de Western blottingAo todo, 40 animais foram usados para os experimentos: 20 eram animais controle e 20 submetidos a tratamento com salicilato. Lâminas com secções frescas (90μm) do tronco cerebral com NCVp, NCD e CI foram obtidas em criostato. Para fazer a biopsia da área desejada, usamos um bisturi circular para retirada de material (punch) de 0,5mm de diâmetro. Devido a seus pequenos tamanhos e localização contígua, o NCD e NCVp foram extraídos no mesmo bisturi. Uma biópsia foi obtida de cada um dos quatro animais e os materiais retirados foram agrupados para aumentar a quantidade de proteína. O tecido foi homogeneizado em tampão de lise com 137mM de NaCl; 20 de mM Tris; Igepal CA‐630 a 1%; glicerol a 10%; 2mM de ortovanadato de sódio; sulfato de dodecilo de sódio a 1%; 50mM de fluoreto de sódio; 2mM de EDTA; e coquetel de inibidores de protease a 10% em pH de 7,4. Os homogeneizados de tecido foram centrifugados a 15.000rpm por 10 minutos a 4°C. A concentração de proteína nos tecidos homogeneizados foi determinada com o uso de um ensaio de Lowry modificado (DC Protein Assay, Bio‐Rad). Alíquotas com 90μg de proteína foram dissolvidas em tampão de carga e aquecidas a 95°C por cinco minutos e as proteínas foram separadas por Tris‐glicina SDS‐PAGE a 7,5% (GE Healthcare‐Bioscience) e transferidas para membranas de PVDF (Amersham Biosciences).
As imunotransferências foram bloqueadas com albumina a 5% e incubadas com anticorpos primários a 4°C. Os anticorpos primários incluíram anticorpos policlonais contra c‐fos e egr‐1 (1:1.000; Santa Cruz Biotechnology). Após a incubação, a membrana foi lavada e incubada com anticorpos secundários (1:10.000; ECL anti‐Rabbit IgG; GE Healthcare) por uma hora em temperatura ambiente. Após as lavagens finais, as proteínas marcadas foram detectadas por quimioluminescência (RPN2132; GE Healthcare). Para a stripping e reprobing, a membrana foi submersa em tampão de extração (100mM de 20‐mercaptoetanol; sulfato de dodecilo de sódio a 2%; 62,5mM Tris‐HCl; pH de 6,8) a 50°C, por oito minutos, lavada por 90 minutos sob água de torneira, reidratada com metanol e lavada com TBS‐T antes de bloquear e ressondar com anticorpos primários contra GAPDH (1:5.000; Abcam). As amostras foram fotografadas e quantificadas com análise de imagem (Molecular Imaging Systems).
Imunoistoquímica de c‐fosDez ratos foram usados, cinco controle e cinco submetidos a tratamento com salicilato. Os ratos foram perfundidos transcardialmente com NaCl a 0,9%, seguido de paraformaldeído a 4% em 0,1M de tampão de fosfato (pH de 7,4). O cérebro foi removido, pós‐fixado na mesma solução por uma hora e crioprotegido com sacarose a 30% em 0,1M de tampão de fosfato por dois dias a 4°C. Os cérebros foram individualmente seccionados (50μM) no plano transversal com um criostato.
As secções foram lavadas em PBS (0,01M, pH de 7,4) e incubadas em PBS com peróxido de hidrogênio a 1% por 30 minutos, para inativar as peroxidases endógenas. Após várias lavagens em PBS por 30 minutos, as secções foram colocadas em soro de cabra normal a 5% (Vector) por uma hora e incubadas por 48 horas a 4°C com anticorpos primários anti c‐fos gerados em coelhos (1:100; Santa Cruz). Depois de lavadas em PBS, as secções foram incubadas por 1h30 à temperatura ambiente com IgG anticoelho de cabra biotinilada (1: 150; Vector). Subsequentemente, foram lavadas em PBS e colocadas por 1h30 em complexo de peroxidase avidina‐biotina (Vectastain, Vector). A imunomarcação foi revelada por incubação de 5 a 10 minutos com tetracloreto de 3,3’‐diaminobenzidina a 0,05% e peróxido de hidrogênio a 0,1%. As secções foram montadas em lâminas revestidas com gelatina, desidratadas, depuradas com xileno e cobertas com laminulas com Entellan®.
Análise dos dadosOs dados da técnica de análise Western blot são apresentados como a razão entre c‐fos e a intensidade do sinal da proteína GAPDH. Os núcleos corados nas seções do CI foram manualmente contados. No total, 11 secções com o CI, nove com o NCD e seis com o NCVp foram contadas para cada animal. Todos os dados foram expressos como média±DP. A análise estatística foi feita com o teste t de Student. Um valor p<0,05 foi considerado estatisticamente significativo.
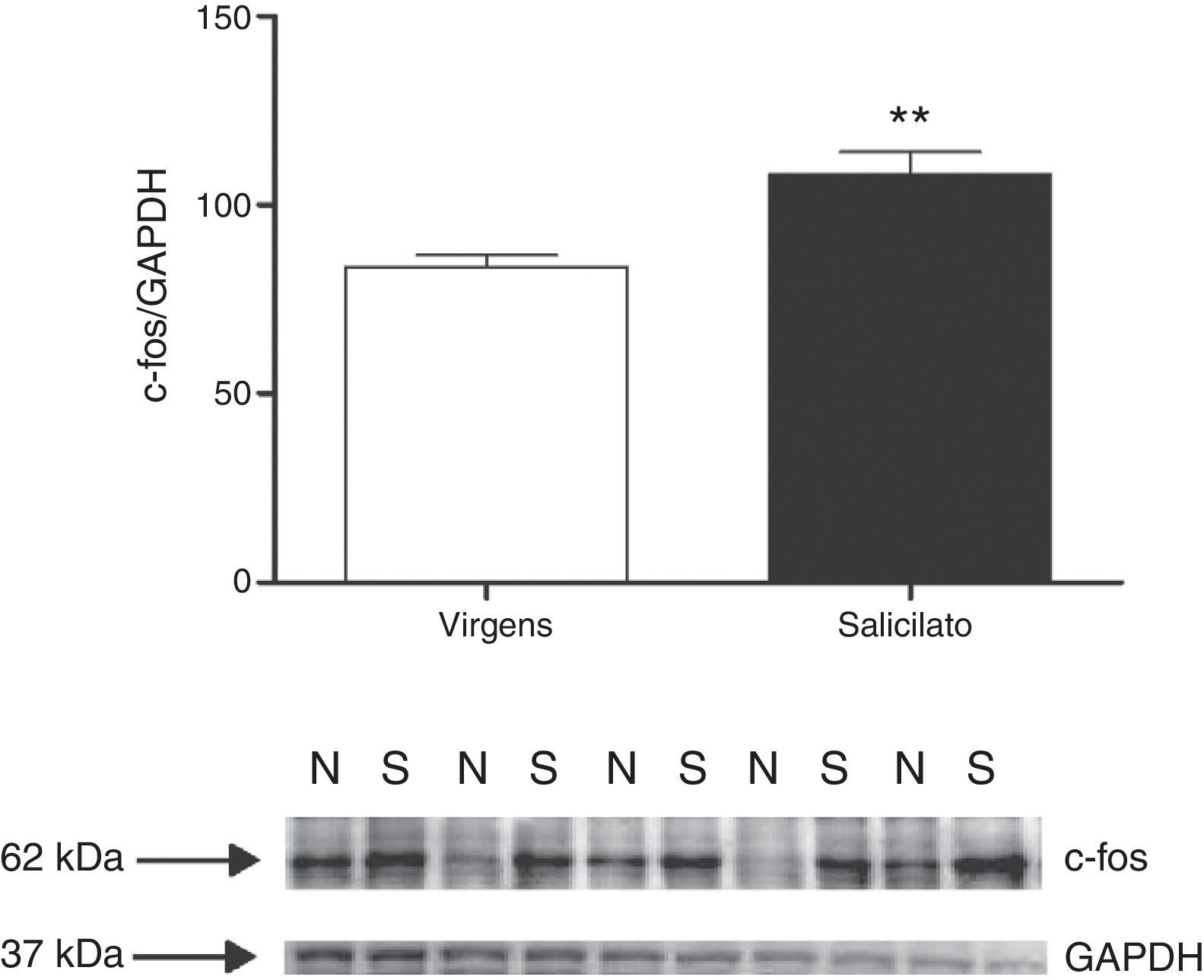
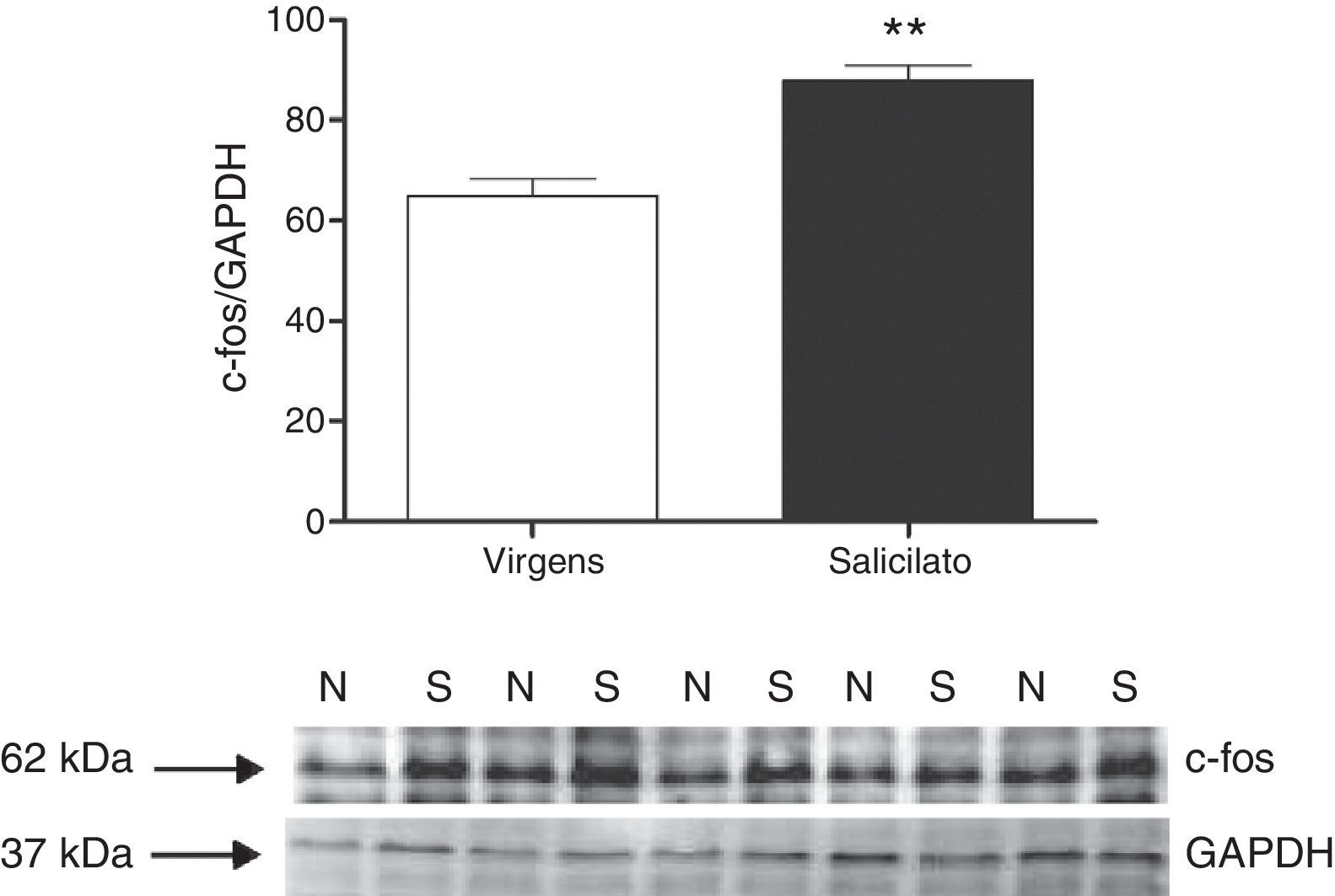
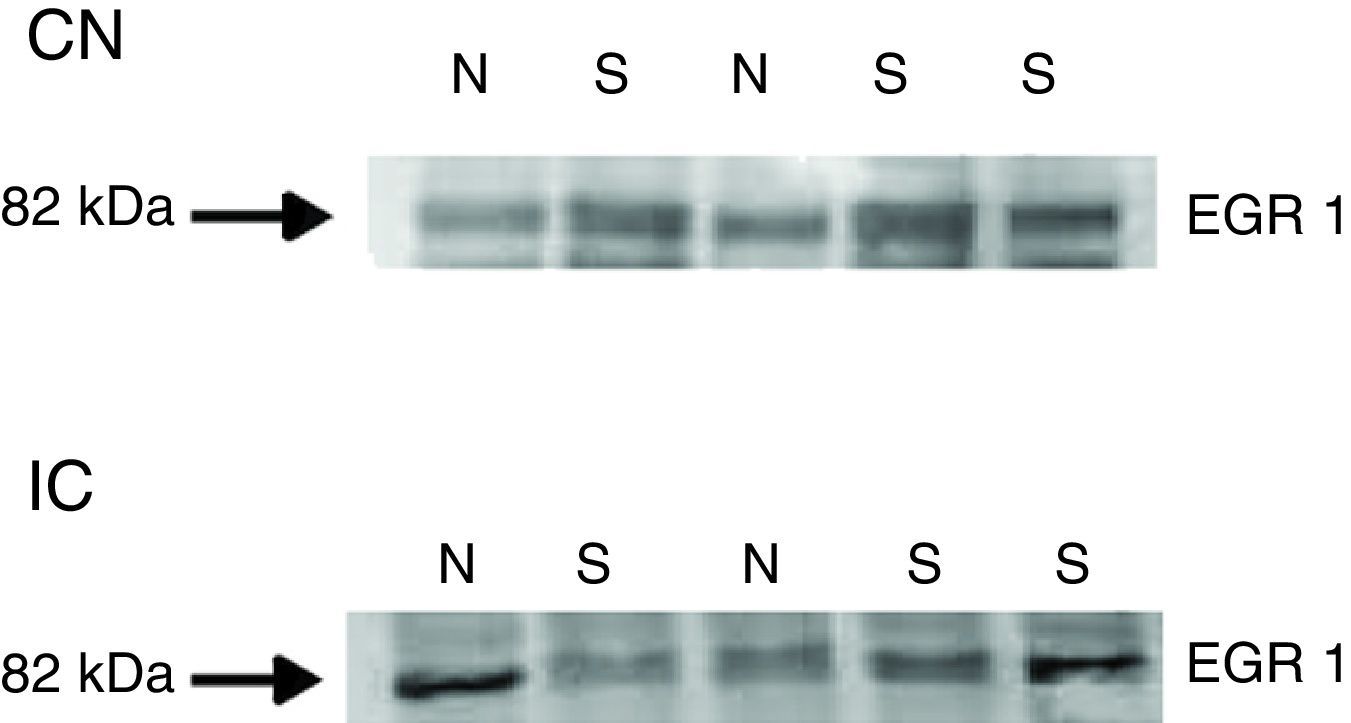
ResultadosApós três dias de tratamento com salicilato (300mg/kg), fizemos as biópsias do tronco cerebral contendo tanto o CI quanto o NCD e o NCVp para análise por Western blot da expressão da proteína c‐fos. Em ambas as áreas, a proteína c‐fos estava significativamente aumentada após o tratamento com salicilato (figs. 1 e 2). Em contrapartida, o aumento da expressão da proteína egr‐1 não foi detectado nessas regiões após o tratamento com salicilato (fig. 3).
. Western blost mostra a imunorreatividade de c‐fos e GAPDH em biópsias (punches) com o NC (N, virgens; S, salicilato).")
Expressão de c‐fos no NC. Superior: quantificação da expressão de c‐fos. A intensidade do sinal de c‐fos foi normalizada com a intensidade de sinal de GAPDH (**p=0,0058; n=5 para cada grupo com material de quatro animais). Western blost mostra a imunorreatividade de c‐fos e GAPDH em biópsias (punches) com o NC (N, virgens; S, salicilato).
. Western blost mostra a imunorreatividade de c‐fos e GAPDH em biópsias (punches) contendo o IC (N, virgens; S, salicilato).")
Expressão de c‐fos no CI. Superior: quantificação da expressão de c‐fos. A intensidade do sinal de c‐fos foi normalizada com a intensidade do sinal de GAPDH (**p=0,0012; n=5 para cada grupo com material de quatro animais). Western blost mostra a imunorreatividade de c‐fos e GAPDH em biópsias (punches) contendo o IC (N, virgens; S, salicilato).
 com o NCD/NCVp e CI de ratos virgens de tratamento e após o tratamento com salicilato de sódio (N, virgens; S, salicilato).")
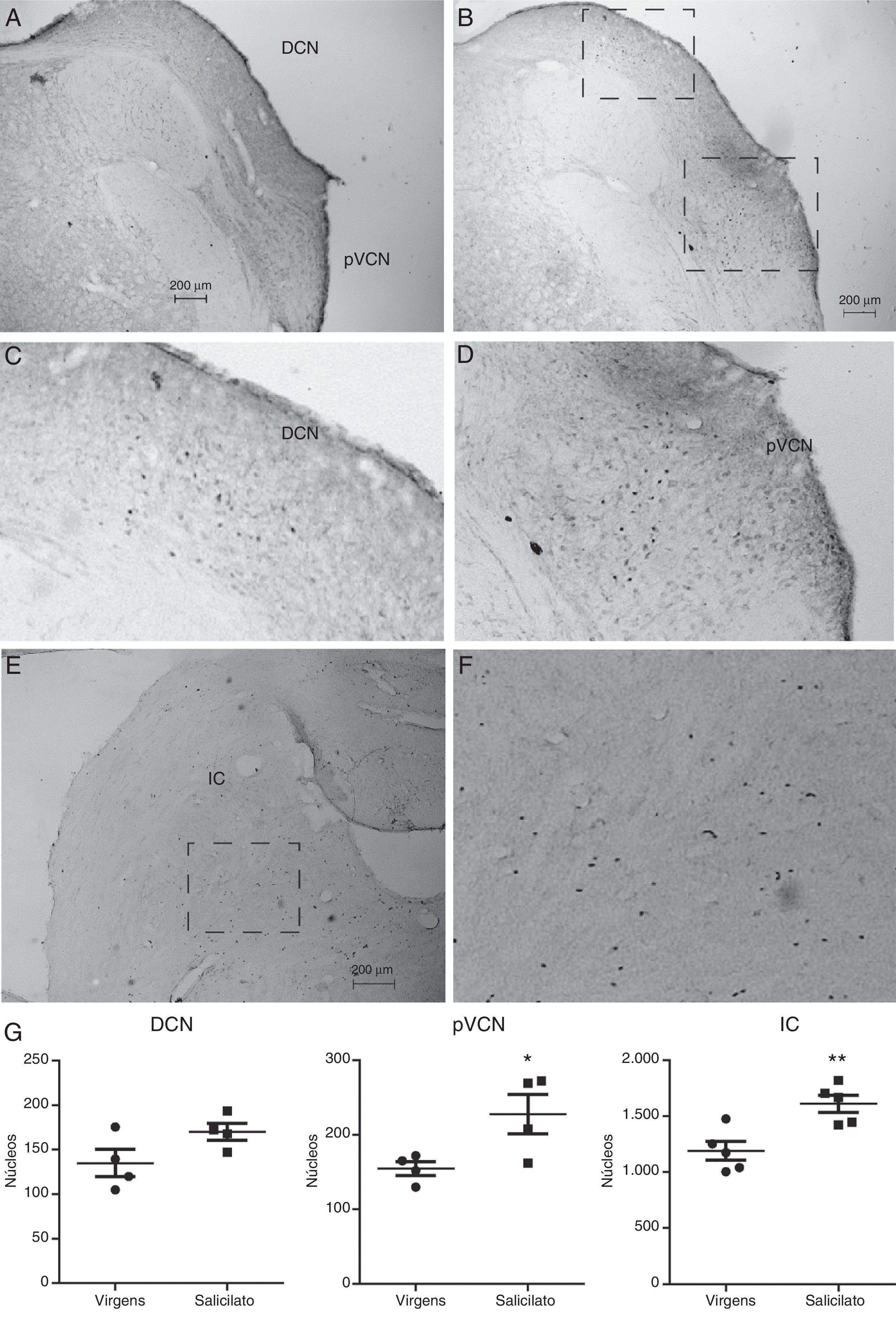
Como a localização da proteína de c‐fos não pôde ser identificada pelo Western blot, fizemos a imunocitoquímica de c‐fos em secções com NCD, NCVp e CI. Detectamos c‐fos em secções de NCD, NCVp e CI, como mostrado na figura 4. Nas secções dos animais virgens de tratamento, uma fraca coloração dos núcleos foi observada em todas as três regiões (fig. 4A). Nas seções dos animais tratados com salicilato, núcleos mais intensamente corados em NCD, NCVp e CI foram observados (fig. 4B). No NCD, essa marcação mais intensa foi observada nas camadas mais internas, camada profunda e possivelmente a camada fusiforme (fig. 4B e C) e observada principalmente na secção medial/dorsal do NCD. Não observamos marcação na camada molecular. Também observamos aumento da rotulagem de c‐fos no NCVp adjacente ao NCD (fig. 4B e D). No CI, observamos um aumento da imunorreatividade, principalmente na parte central, como pode ser visto na figura 4E e 4F. Não observamos rotulagem na oliva superior (não mostrado).
![Detecção de c‐fos por imunocitoquímica no NCD e CI. A, Secção com o NCD e o núcleo coclear ventral posterior (NCVp) de um animal virgem de tratamento. B, Secção com o NCD e o NCVp de um animal tratado com salicilato. C, Detalhe dos núcleos que expressam c‐fos no NCD a partir do quadrado mostrado em B. D, Detalhe dos núcleos que expressam c‐fos no NCVP a partir do quadrado mostrado em B. E, Seção com o CI de um animal tratado com salicilato. F, Detalhe de núcleos que expressam c‐fos no CI a partir do quadrado mostrado em E. G, Quantificação da expressão de c‐fos no NCD, NCVp e CI [*p=0,04; **p=0,006; n=4 (NCD, NCVP) e 5 (CI)].](https://static.elsevier.es/multimedia/25300539/0000008300000002/v1_201703310220/S253005391730007X/v1_201703310220/pt/main.assets/gr4.jpeg?xkr=ue/ImdikoIMrsJoerZ+w9xMbaXYzIj2UVZ9WuQ9rDVcCk/jSInH8zfFEkCTBR2+UewNUNhDYczZ4+mvtXdqpisGaQOudDkpr86ie6IqKlN8VLop3ZyXR/uVj4GJVO2bwaIFe2w5WjdMVY4TPbvHY/6k11TuZWIJFTvtQtQJahww65v/IX+0GhwgHAn6RfNyDAUI7AJadEb75ZqGkYXyuqrj7tPQ06Y6Lg0cMEBF3bV2ws6kA+ar17iYIT6bk+Nhvfiz0GNn2PiYNWc626bAWkCCgbfJDZ7FdsWs/Oy0kKIXzev8WC5B2iIP4YhIprML2 "Detecção de c‐fos por imunocitoquímica no NCD e CI. A, Secção com o NCD e o núcleo coclear ventral posterior (NCVp) de um animal virgem de tratamento. B, Secção com o NCD e o NCVp de um animal tratado com salicilato. C, Detalhe dos núcleos que expressam c‐fos no NCD a partir do quadrado mostrado em B. D, Detalhe dos núcleos que expressam c‐fos no NCVP a partir do quadrado mostrado em B. E, Seção com o CI de um animal tratado com salicilato. F, Detalhe de núcleos que expressam c‐fos no CI a partir do quadrado mostrado em E. G, Quantificação da expressão de c‐fos no NCD, NCVp e CI [*p=0,04; **p=0,006; n=4 (NCD, NCVP) e 5 (CI)].")
Detecção de c‐fos por imunocitoquímica no NCD e CI. A, Secção com o NCD e o núcleo coclear ventral posterior (NCVp) de um animal virgem de tratamento. B, Secção com o NCD e o NCVp de um animal tratado com salicilato. C, Detalhe dos núcleos que expressam c‐fos no NCD a partir do quadrado mostrado em B. D, Detalhe dos núcleos que expressam c‐fos no NCVP a partir do quadrado mostrado em B. E, Seção com o CI de um animal tratado com salicilato. F, Detalhe de núcleos que expressam c‐fos no CI a partir do quadrado mostrado em E. G, Quantificação da expressão de c‐fos no NCD, NCVp e CI [*p=0,04; **p=0,006; n=4 (NCD, NCVP) e 5 (CI)].
A análise quantitativa dos núcleos que expressam c‐fos demonstrou um aumento significativo de núcleos positivos para c‐fos no NCVp (controle: 155±9,2 núcleos; salicilato: 228±26,4 núcleos; p=0,04, n=4) e no CI (controle: 1190±86 núcleos; salicilato: 1611±79 núcleos; p=0,0061, n=5), mas não no NCD (controle: 135±15 núcleos; salicilato: 170±9,5 núcleos; p=0,1; n=4), como mostrado na figura 4G.
DiscussãoNossos dados mostram que o tratamento por três dias com doses elevadas de salicilato aumenta a expressão do produto do GPI c‐fos, mas não de egr‐1 no NCVp, NCD e CI de ratos. Embora tenhamos observado o aumento dos níveis da proteína c‐fos e uma marcação nuclear mais intensa no CI, NCD e NCVp após o salicilato, o número de núcleos corados foi significativamente maior apenas no NCVp CI, mas não no NCD. Esses dados estão de acordo com a hiperatividade de NCD e CI observada em ratos submetidos a um protocolo similar de tratamento com salicilato3,15,16 e em outros modelos animais de zumbido.17,18 Por outro lado, observamos um aumento significativo na expressão de c‐fos no NCVp adjacente, uma região que geralmente não está relacionada à indução de zumbido.3 Esse aumento da expressão de c‐fos no NCD, NCVp e CI sugere que a expressão desses GPIs promove a expressão de proteínas que podem alterar as propriedades fisiológicas e morfológicas dos neurônios dessas regiões, o que poderia ser um substrato para a indução de zumbido.
Outros pesquisadores4–7 avaliaram a expressão de c‐fos após o tratamento agudo com salicilato com o uso de imunocitoquímica. Esses estudos revelaram que a expressão de c‐fos não aumentou consistentemente no NCD e CI. Nossos resultados, por outro lado, mostram uma clara expressão aumentada de núcleos positivos para c‐fos em CI e NCVp. Acreditamos que essas discrepâncias podem ser atribuídas ao fato de que usamos animais mais jovens e que esses estudos administraram uma dose única de salicilato de sódio, em vez das três doses diárias administradas por nós. Consistente com essa hipótese, um estudo que usou seis dias de tratamento com salicilato também observou um aumento significativo de núcleos positivos para c‐fos no CI, mas não no NCD.8
Em animais experimentais, o tratamento com salicilato aumenta a descarga de neurônios em ambos NCD e CI.15,16 Os mecanismos desse efeito ainda são desconhecidos e poderiam refletir um efeito direto do salicilato na excitabilidade neuronal desses neurônios ou de suas próximas sinapses, ou um efeito da expressão dos GPIs induzido pelo salicilato. Ao contrário de um efeito excitatório direto do salicilato, demonstrou‐se que a aplicação direta de salicilato em neurônios do NCD em fatias de cérebro diminui a descarga espontânea e evocada dos principais neurônios do NCD.19,20 Por outro lado, a aplicação direta de salicilato aumenta a descarga da maioria dos neurônios do CI in vitro.21 O salicilato também poderia aumentar a excitabilidade dos neurônios de NCD e CI e diminuir suas transmissões inibitórias glicinérgicas.22 Porém, as correntes glicinérgicas em neurônios fusiformes do NCD não foram afetadas pelas aplicações agudas de concentrações milimolares de salicilato.20 O salicilato também pode inibir os receptores GABAérgicos23 e aumentar as correntes de NMDA,24 o que poderia potencialmente perturbar o equilíbrio entre excitação‐inibição. Como a maioria dos neurônios do NCD é espontaneamente disparada,25 um aumento da descarga produzido por salicilato pode não aumentar de forma significativa o número de neurônios expressando c‐fos, o que poderia explicar o efeito não significativo do salicilato no número de núcleos marcados com c‐fos no NCD. Por outro lado, nossos dados mostraram um aumento significativo na expressão de núcleos de c‐fos no NCVp. Isso sugere que o salicilato ativava seus neurônios e que essa região poderia ser relevante para a percepção de zumbido.
Tanto c‐fos quanto egr‐1 são expressos nas vias auditivas em resposta à estimulação sonora e intracoclear.10,12,26,27 Curiosamente, a expressão de c‐fos no NCD provocada pela estimulação intracoclear foi mais observada em interneurônios glicinérgicos na camada molecular.27 Essa descoberta é intrigante, uma vez que esses interneurônios não recebem input da via auditiva primária, mas das fibras paralelas que transmitem principalmente informações somatossensoriais. A marcação de c‐fos foi observada apenas perto ou na camada profunda do NCD em ambos os grupos controle e salicilato. Isso sugere que c‐fos é expresso principalmente pelos neurônios que computam a informação auditiva. Por outro lado, não encontramos aumento de expressão de egr‐1, um GPI que está ligado à formação de plasticidade em longo prazo,13 após o tratamento com salicilato. Isso sugere que o salicilato não desencadeia a expressão de genes relacionados com a plasticidade sináptica por muito tempo. Curiosamente, um estudo anterior observou uma diminuição da expressão do gene egr‐1 no CI após o tratamento com salicilato.28
Há semelhanças entre o zumbido e a dor neuropática.29,30 Isso sugere que podem compartilhar mecanismos moleculares semelhantes. Por exemplo, os pacientes com zumbido muitas vezes apresentam hiperacusia e os pacientes com dor crônica apresentam hiperalgesia, ambas reações exacerbadas à intensidade do estímulo normal. Como proposto para o zumbido,31 acredita‐se que a dor crônica seja um resultado de alterações plásticas em longo prazo nos neurônios sensoriais e sinapses.32 O GPI c‐fos também é ativado na medula espinhal em modelos de dor crônica e inflamação.33,34 Sua expressão é postulada como o primeiro sinal de alterações nesses neurônios que levará à dor crônica. Portanto, é provável que c‐fos ative cascatas semelhantes de expressão do gene no zumbido e na dor crônica, o que leva a alterações que seriam subjacentes à sintomatologia dessas condições.
ConclusãoNossos dados mostram que as doses de salicilato indutoras de zumbido3 aumentam a expressão de c‐fos, mas não de egr‐1 em áreas auditivas do tronco cerebral: núcleo coclear dorsal, núcleo coclear posteroventral e colículo inferior de ratos. Portanto, postulamos que c‐fos ativa cascatas de expressão do gene nessas áreas, o que leva a alterações que poderiam ser a base da sintomatologia do zumbido.
Conflitos de interesseOs autores declaram não haver conflitos de interesse.
A revisão por pares é da responsabilidade da Associação Brasileira de Otorrinolaringologia e Cirurgia Cérvico‐Facial.
Como citar este artigo: Santos P, da Silva LE, Leão RM. Specific immediate early gene expression induced by high doses of salicylate in the cochlear nucleus and inferior colliculus of the rat. Braz J Otorhinolaryngol. 2017;83:155–61.
gology tem o prazer em homenagear os revisores


